这些问题将通过回顾(非水族馆)实地研究的相关文献以及对从一系列水族馆水族箱中采集的海洋水族馆水样进行实验来探讨:
简介: DOC、TOC、珊 瑚细菌和珊 瑚健康之间的合理联系由于捕鱼、旅游和海岸保护,全世界珊 瑚礁的经济价值估计为每年 375 亿美元(Costanza,1997 年)。不幸的是,在世界许多地区,珊 瑚礁正在死亡;例如,加勒比珊 瑚礁在过去 30 年中遭受了大约 80% 的珊 瑚种群损失(Gardner,2003 年)。这些相互依存的事实激发了许多科学领域的努力,既要确定珊 瑚(=珊 瑚礁)死亡的原因,又要寻求可能的改善干预措施。许多人为(=归因于人类活动)入侵珊 瑚礁生态系统已被认为是造成热带珊 瑚礁上珊 瑚的压力甚至死亡的原因,尽管也不能排除自然发生的压力源。所有这些活动都引起了人们对造礁珊 瑚的生态意义及其对环境变化的反应的关注。由于饲养实践中的计划外和不受欢迎的偏差,许多对热带珊 瑚礁的珊 瑚窘迫/死亡率的观察可以在我们的(压力)水族馆中重复。因此,保持健康并确保我们在家庭水族箱中精心培育的脆弱生物的生存能力仍然是一项重大挑战,尤其是在水箱老化和废物积累的情况下。由于饲养实践中的计划外和不受欢迎的偏差,许多对热带珊 瑚礁的珊 瑚窘迫/死亡率的观察可以在我们的(压力)水族馆中重复。因此,保持健康并确保我们在家庭水族箱中精心培育的脆弱生物的生存能力仍然是一项重大挑战,尤其是在水箱老化和废物积累的情况下。由于饲养实践中的计划外和不受欢迎的偏差,许多对热带珊 瑚礁的珊 瑚窘迫/死亡率的观察可以在我们的(压力)水族馆中重复。因此,保持健康并确保我们在家庭水族箱中精心培育的脆弱生物的生存能力仍然是一项重大挑战,尤其是在水箱老化和废物积累的情况下。
幸运的是,有大量的信息,有些是轶事,有些是经过严格的实验验证的,有责任心的鱼友可以帮助完成这项任务。许多(大多数?)与成功饲养相关的规范参数已被确定。照明、流量、温度、进料等问题通常可以通过几种不同且有效的方法来解决。维持水的化学性质可能比较棘手,因为必须平衡关键和微量水成分的消耗/去除和生产/添加。然而,主要元素/化合物的可接受范围已记录在案,监测和维持适当水平的盐度、碱度、钙、镁、锶、碘化物、硝酸盐、氨、磷酸盐等所需的方法是众所周知且易于获得的.
什么是溶解有机碳 (DOC)、颗粒有机碳 (POC) 和总有机碳 TOC)?在这种情况下,碳是一个笼统的短语,指的是含碳(即有机)化学物质,它们要么
溶解在水中(= 溶解有机碳,DOC),要么作为小颗粒悬浮在水中,包括单细胞生物(= 颗粒有机碳,POC)(Holmes-Farley,2004)。这些碳源统称为总有机碳,TOC。DOC 和 POC 之间的划分是任意的,并且严格基于市售过滤材料的能力。目前,DOC的操作定义包括通过0.2微米过滤器孔隙的含碳材料。任何留下的含碳材料,包括大多数细菌/单细胞生物,都被标记为颗粒有机碳 (POC) (Benner, 2002)。
广告
颗粒和溶解形式的有机碳被称为“海洋土壤”,以表彰其在整个海洋生态系统的基础上所发挥的作用(Ducklow,2002 年)。它是海洋中微生物群的维持,整个海洋食物金字塔都依赖于这种燃料来源的可用性。海洋中超过 97% 的有机物以 DOC 的形式存在,尝试量化所有海洋水中的 DOC 含量会得出惊人的值:650 – 700 x 10 15克 C,一个数字与大气中的碳总量(以 CO 2计)相同数量级(~ 750 x 10 15克)(Hedges,2002;Benner,2002)。
由于在极其复杂的相似物种混合物中分离和表征极少量离散分子的困难,这种聚合 DOC 的化学组成在很大程度上仍然是一个谜。尽管如此,在现代分离(色谱)技术和复杂的表征方法(核磁共振、质谱)的帮助下,已经取得了一些进展(Benner,2002)。已确定了广泛类别的生物代谢物成员,对 DOC 组成的最佳估计值徘徊在以下值附近:
- ~ 6% 的 DOC 来自可水解糖,包括葡萄糖、半乳糖、岩藻糖、甘露糖和木糖。
- ~ 3% 来自可水解氨基酸,包括甘氨酸、精氨酸、丙氨酸、苯丙氨酸、丝氨酸、苏氨酸、天冬氨酸、缬氨酸、组氨酸、异亮氨酸、谷氨酸、赖氨酸、蛋氨酸和亮氨酸。
- ~ 1% 来自可水解的氨基糖,包括N-乙酰氨基葡萄糖和N-乙酰半乳糖胺。
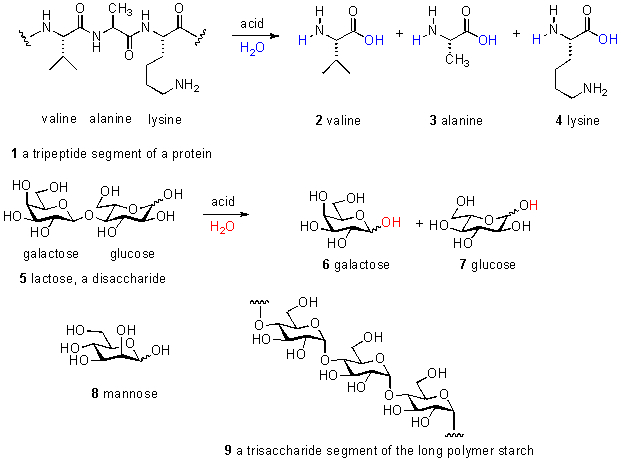
因此,这些常见的生化物种只能占总 DOC 的 10%;其余 90% 的 DOC 仍未定性。这种未知的大部分碳被称为“海洋腐殖质”或“耐火碳”,但这些标签并没有提供有用的化学信息。事实上,即使声称上面列出的碳水化合物和氨基酸存在于 DOC 中也是一种误导;这些物种实际上主要作为聚合物存在,其中许多氨基酸在称为肽或蛋白质的链中连接在一起,并且许多单独的糖分子同样在称为寡糖的(有时是支链的)链中连接在一起。只有通过酸催化水解(=水的化学加成),这些聚合物才能分解成它们各自的组分并如上所述进行表征。这些物种作为单个单元和聚合物链的一部分的一些例子如图 1 所示。海洋中的大部分 DOC 在数小时到数周的时间跨度内被消耗。其余的最终定居在深海中,14 C(= 放射性碳)测年技术已将其年龄置于 4000 – 6000 年范围内(Benner,2002)。
检查图 1 所示的化学结构强调了一个事实,即构建生命分子还需要许多其他元素。已经测量/计算了不同生物的碳百分比(按干重计),一些示例的可靠值是:底栖无脊椎动物 30 – 49% C;中上层无脊椎动物 45-50% C; 人类 28% C (Liess, 2006; Sterner, 2002)。因此,当在测定中报告 1 ppm 的 C 值时,有机材料的实际量在 2 到 3.3 ppm 之间(按质量计)。
热带珊 瑚礁水域中碳(溶解和/或颗粒)的自然水平是多少?在深海?海洋中有机碳水平(DOC 和 POC)的测量有着悠久而丰富多彩的历史。Natterer 首次发表的尝试可以追溯到 1892 年,自那次披露以来,该主题一直停留在化学海洋学家的视野中。检测浸入盐和无机碳“海洋”中的少量碳(碳酸氢盐,HCO 3 –) 已被证明是一个令人烦恼的挑战,并且已经探索了许多方法。直到 1990 年代,当前的“最佳”方法才作为商业仪器可用性的结果出现,尽管关键仪器的“自制”版本可以追溯到 1960 年代。这种称为高温氧化燃烧 (HTOC) 的方法实际上是燃烧样品中的所有有机残留物以从碳中产生 CO 2,然后可以使用适当的检测器识别和量化新形成的 CO 2 。即使有了这样的进步,高问题
仪器“空白”困扰着早期获取海洋 DOC 水平有用数据的尝试,但 1991 年的一次会议和随后的后续行动导致了有关样品制备和适当空白利用的方法的规范化(Sharp,1993;Sharp, 2002)。
选择的商业仪器是 Shimadzu 5000 TOC 分析仪。确定仪器详细信息后,记录了珊 瑚礁和公海中 DOC、POC 和 TOC 含量的许多可靠测量值,表 1。深海 TOC(在本例中为 DOC)的普遍接受值范围为 ~ 0.45 – 0.60 ppm,这个数字似乎对收集位置不敏感。然而,在珊 瑚礁上,DOC(和 TOC)值要高得多。即使注意到这一点,DOC 对从南太平洋到日本到加勒比海到
红海的珊 瑚礁的价值在它们的范围内非常一致:0.7 – 1.6 ppm。波纳佩岛一些地点的异常高值归因于陆地径流(=污染),类似于东京湾和佛罗里达湾的高值。因此,在健康繁荣的珊 瑚礁的散装水中似乎有一个共识值为 1.1 ± 0.4 ppm 的碳。这些珊 瑚礁,至少在南太平洋,只是栖息着构成我们水族馆生物群的珊 瑚、鱼类和其他牲畜的生态系统类型。
| 入口 | 地点 | 深度(英尺) | 有机溶剂(ppm) | POC(ppm) | 总有机碳(ppm) | 参考。 |
| 1 | Haraiki 泻湖,FP a | 27 | 0.76 | 0.17 | 0.93 | 佩吉斯 (1997) |
| 2 | Hikueru泻湖,FP | 58 | 0.93 | 0.11 | 1.04 | 佩吉斯 (1997) |
| 3 | 希提泻湖,FP | 26 | 0.92 | 0.12 | 1.04 | 佩吉斯 (1997) |
| 4 | Kauehi泻湖,FP | 104 | 0.82 | 0.09 | 0.91 | 佩吉斯 (1997) |
| 5 | Marokau 泻湖,FP | 65 | 0.85 | 0.11 | 0.96 | 佩吉斯 (1997) |
| 6 | Nihiru泻湖,FP | 58 | 0.72 | 0.11 | 0.73 | 佩吉斯 (1997) |
| 7 | Rekareka 泻湖,FP | 3 | 1.00 | 0.22 | 1.22 | 佩吉斯 (1997) |
| 8 | Taiaro 泻湖,FP | 39 | 1.67 | 0.19 | 1.86 | 佩吉斯 (1997) |
| 9 | Tekokota 泻湖,FP | 10 | 0.72 | 0.09 | 0.81 | 佩吉斯 (1997) |
| 10 | Tepoto Sud 泻湖,FP | 16 | 0.74 | 0.12 | 0.86 | 佩吉斯 (1997) |
| 11 | 马歇尔群岛马杰罗环礁。 | <1 | 0.62-2.18 ℃ | | | 吉永 (1991) |
| 12 | 波纳佩岛,密克罗尼西亚 | <1 | 1.61-4.51 ℃ | | | 吉永 (1991) |
| 13 | 大星盘泻湖,斐济 | 65-120 | 1.37 | | | 托雷顿 (1999) |
| 14 | 日本白穗礁 | <1 | 0.86 | 0.04 | 0.90 | 羽田 (2002) |
| 15 | 库拉索,北美b(天) | 23 | 1.55天 | | | 范杜伊尔 (2001) |
| 16 | 库拉索,NA(夜) | 23 | 1.16天 | | | 范杜伊尔 (2001) |
| 17 | 珊 瑚礁 Nat。水库,以色列 | 29 | 0.86 | 0.09 | 0.95 | 亚赫尔 (2003) |
| 18 | 东京湾 | <1 | 1.92 | | | 小川 (1992) |
| 19 | 开阔的海洋,马杰罗环礁 | 1-162 | 0.88-1.66 ℃ | | | 吉永 (1991) |
| 20 | 公海等 太平洋 | <1 | | < 0.06 | 0.84 | 夏普 (1995) |
| 21 | 公海等 太平洋 | 13000 | | < 0.01 | 0.47 | 夏普 (1995) |
| 22 | 开阔的海洋,靠近 FP | — | 1.00 | 0.05 | 1.05 | 佩吉斯 (1997) |
| 23 | 开阔的海洋,马尾藻海 | 6 | 0.88 | | | 夏普 (1993) |
| 24 | 开阔的海洋,马尾藻海 | 7150 | 0.59 | | | 夏普 (1993) |
| 25 | 佛罗里达州佛罗里达湾 | <1 | | | 10.5 | 博耶 (1997) |
| 法属波利尼西亚。b荷属安的列斯群岛。c多个收集点的范围。d湿化学氧化法,与 HTOC 法互相关。 |
在珊 瑚礁环境中,哪些生物产生碳,哪些生物消耗碳?珊 瑚礁环境中 DOC 的主要消费者是异养(= 不能自己制造食物)细菌(Moriarty,1985 年)。这些细菌可以生活在
水柱中或与底物、碎屑、珊 瑚和其他底栖生物等紧密相关。细菌是珊 瑚礁食物网中的关键组成部分,因为它们在从源头(=阳光)转移能量中扮演“中间人”的角色) 给珊 瑚礁上的消费者(Eppley,1980;Ducklow,1983;Crossland,1980;Johannes,1967;Ducklow,1979;Gottfried,1983)。照射在珊 瑚礁上的阳光通过促进水柱中自由游动的浮游生物以及我们在水族馆中痴迷的石珊 瑚虫黄藻内的光合作用来启动能量循环。光合作用利用阳光的能量将无机碳以 CO 2的形式“固定”(= 附着)到最终变成碳水化合物的有机化学结构上(参见图 1)。这些
碳水化合物结构单元进一步由虫黄藻和/或珊 瑚宿主进行化学操作,然后由珊 瑚分泌为珊 瑚粘液(复杂多糖、氨基酸低聚物/聚合物、脂质等的组合,参见图 1)。粘液中的碳水化合物和氨基酸是构成海洋食物金字塔基础的细菌和其他微生物群的食物来源(Kirchman,1990;Rich,1996;Weiss,1999;Wild,2004;Sharon,2008)。剩下的珊 瑚礁居民,包括居住着虫黄藻的珊 瑚,然后以这些富含能量的微生物为食(或者反过来,以食用微生物的生物为食,无止境),从而使珊 瑚礁的营养循环食物网永久存在。除了珊 瑚,海绵是海洋细菌最多产的储存库之一。实际上,
广告
DOC从哪里来?浮游植物是主要贡献者,并且提出了一个有趣的假设,即当营养水平不足以生长时,释放 DOC 是一种处理过量、光合作用产生的“固定”碳的方法(Hessen,2008 年)。如上所述,珊 瑚礁上 DOC 的一些主要生产者是珊 瑚本身(Crossland,1987)。索罗金使用一些颇具创意的碳平衡核算方法,估计热带珊 瑚礁水域中约 20% 的 DOC 可归因于珊 瑚粘液的产生(Sorokin,1993 年)。在一项独立研究中,珊 瑚礁沉积群落代谢的总 TOC 的相似值(10 – 20%)归因于珊 瑚粘液(Wild,2004 年);Johannes,使用不同的估计方法和在不同的地点,将该值缩放回 ~ 2% (Johannes, 1967)。即使由于所做的假设,这些数字中有很大的误差线,令人惊讶的是,珊 瑚为珊 瑚礁水柱贡献了如此多的有机物质,并且通过推断,对我们(库存过多?)珊 瑚礁水族馆的封闭系统做出了贡献。
其他几条证据支持这样的论点,即珊 瑚通过它们的粘液分泌物,实际上用 DOC 淹没了珊 瑚礁。例如,与附近的珊 瑚礁水(1.60 – 1.94 ppm)相比,在库拉索岛附近的礁石上测量Montastraea faveolata和奇异果树周围接触地表水的 DOC 含量(约 2.28 ppm)(van Duyl,2001 年) )。在另一项(基于实验室的)研究中,观察到束状藻每天早上和每天下午都会发出一个持续约一个小时的 DOC“尖峰”。与未喂食的对照相比,喂食卤虫的标本分泌更大的 DOC 峰值:对于喂食的Galaxea,在 5 升水箱内测得的 DOC 值从 1.2 ppm 的基础值提高到 4.8 – 20.4 ppm;未喂食的 Galaxea在尖峰排放期间仅将水的基础 DOC 水平提高了几个 ppm。在这两种情况下,在峰值后 2 小时内,水库中的 DOC 含量回落至基础水平(Ferrier-Pagès,1998)。碳平衡计算表明,约 11-14% 的光合固定碳(即源自 HCO 3 - <-> CO 2)以 DOC 形式释放,该值与许多先前的研究一致(Sorokin,1993;Crossland,1987 )。
当Galaxea峰值消退时,DOC 去哪里了?14 C 标签研究表明,它会被生活在珊 瑚内和珊 瑚上的细菌迅速消耗,而不是被水柱中的细菌消耗。通过抗生素治疗关闭这些内源性细菌消除了 DOC 的吸收。
在随后的一项研究中,Ferrier-Pagès 及其同事扩展了这种类型的实验,在他们的实验室水族馆中加入了更加丰富多样的珊 瑚画面(Ferrier-Pagès,2000)。Stylophora sp的标本。,鹿角藻。, Favia sp。, Galaxea sp。, 和Euphyllia ancora在一个实验池中,类似地,Montipora sp。, Favia sp。又是安科拉真叶植物在第二个独立水箱中,用微型浮游生物(水箱 1)和海洋细菌(水箱 2)处理。在这两种情况下,在喂食后 2 – 7 小时检测到 DOC 增加了 5 – 13 倍基线。这些 DOC 爆发在大约 2 小时后消退,并且 DOC 水平下降到喂食前的值。在实验过程中,对照水箱(底物但没有珊 瑚)没有表现出任何 DOC 激增。Means 和 Sigleo 可以对Acropora palmata进行类似的观察以及(均值,1986 年)。总之,这些对珊 瑚摄食与随后的 DOC 生产和消费之间相关性的一致观察,在广泛的珊 瑚物种和食物类型中,开始说明这一现象的普遍性。事实上,这些观察结果可能预示着一种假设的发展,即珊 瑚通过以 DOC 的形式计量食物,故意调节其细菌种群的数量和含量(见下文 Rohwer)。
珊 瑚与其相关微生物群之间明显的紧密耦合,可能至少部分由 DOC 的产生/消耗介导,这表明这种细菌种群的整体“健康”可能会影响珊 瑚本身的整体健康。这一概念已被罗森伯格扩展和编纂为“进化的全基因组理论”,该理论规定宿主生物(即珊 瑚)及其相关微生物群(细菌、虫黄藻等)应被视为一个单一的进化单位; 全息生物(罗森伯格,2007)。将细菌的繁殖时间(分钟)与珊 瑚的繁殖时间(月?年?)进行比较,可以看出这种观点的价值。与宿主珊 瑚相比,微生物共生体的遗传信息可以通过突变和/或水平基因转移更快地改变,从而确保全生物体可以通过自然选择更快地适应快速变化的环境条件。为了支持这一概念,珊 瑚粘液层中测量的细菌数量是周围水中的 100 - 1000 倍。此外,LaJeunesse 的工作表明,宿主珊 瑚物种与与之相关的虫黄藻(= 共生藻类)的特定进化枝之间存在相关性,并且这些相关的关联对进化压力有反应(LaJeunesse,2005a;LaJeunesse,2005b;另见 Baker,2003)。同样,在不同地理位置的同种珊 瑚中发现了相似的细菌种群组成,而不同物种的并列珊 瑚对其全息生物具有很大不同的细菌成分(Rohwer,2001,2002;Bourne,2005;Rosenberg,2007;Ritchie, 2008)。或者,一些证据表明环境梯度(例如深度、污染)是细菌种群的决定因素(Klaus,2007)。因此,尽管珊 瑚/细菌关联的最终图景无疑将是复杂和多因素的,但似乎有某种程度的持久珊 瑚-微生物选择性表明共生体与宿主有目的地匹配。我们水族馆中的珊 瑚及其相关微生物群对珊 瑚礁研究的珊 瑚有类似的反应吗?初步证据表明,事实上,相同的微生物种群粒状真菌水族馆和野外的珊 瑚不同,足以引起人们的兴趣(Kooperman,2007)。作者认为,水族馆标本与珊 瑚礁标本中微生物群的差异是珊 瑚全息生物对其环境适应性的一个例子。有趣的是推测珊 瑚故意控制它们的细菌种群以赋予生存优势,并且 DOC 分泌可能是实施这种控制的一种机制。关于这一点的一些令人鼓舞的间接证据已经记录在案。例如,不同种类的藻类在成分上贡献不同的 DOC,而这些化学成分的差异显然与不同细菌种类的募集有关(González,2000;Pinhassi,2004;Grossart,2005;相关示例另见 Covert,2001) .假单胞菌科(P) 和交替单胞菌科/ Cotwelliaceae (A/C) 具有定义的各种碳水化合物和氨基酸输入(作为 DOC 组分的模型)导致 P:A/C 比率作为碳源输入函数的显着差异(Allers ,2007a;阿勒斯,2007b)。最后,观察到(1)珊 瑚粘液的化学成分因物种而异(Meikle,1988),(2)水生细菌和珊 瑚细菌的不同遗传变体Alteromonadaceae macleodii与不同的珊 瑚相关(伯恩,2005 年;兰伯特,
2006 年);(1) 是否可以确定性地与 (2) 相关联?



 [/url]
[/url]
 [/url][url=]
[/url][url=] [/url][url=]
[/url][url=] [/url][url=]
[/url][url=] [/url]
[/url] [/url][url=]
[/url][url=] [/url][url=]
[/url][url=] [/url][url=]
[/url][url=] [/url][url=]
[/url][url=] [/url]
[/url]
 [/url][url=]
[/url][url=] [/url][url=]
[/url][url=] [/url][url=]
[/url][url=] [/url]
[/url]




