本帖最后由 飒枫 于 2020-8-9 21:37 编辑
以后这类的系列文章会非常硬核,硬核玩家可以狂喜了。对了硬核玩家们,我已经开了微信公众号了,不想错过任何新文章的硬核玩家们可以关注哦。
对于鱼友来说,最关注的东西大概就是shanhu生长和发色,生长的事情太繁杂散乱了,咱们可以来先说说发色的事情。 发色这个事情也算是智商税重灾区了,这个我已经吐槽过无数次了,毕竟随便拿点各种盐配一配就能卖高价这种事情简直不要太爽。 可是对于真正的荧光蛋白、色蛋白来说,它们与鱼友想象的却大不一样。
归功于荧光蛋白在生物学研究里已经得诺奖的巨大应用,荧光蛋白的结构,光学特征、化学性质等等已经被学界了解的非常清楚了,但是对于荧光蛋白在shanhu体内起到的功能,shanhu对荧光蛋白的调控机理我们所知的却非常少。 为了彻底破除大家对一些补剂、玄学的迷信,为了对科学知识、精神的传播,我就来写一个荧光蛋白、色蛋白的系列科普。
今天第一篇就从荧光蛋白的结构、进化说起。 和大多数鱼友想象的不同,荧光蛋白的种类可以说是五花八门,目前已经确定的荧光蛋白有几百种之多,这个可以参考下我以前写过的shanhu荧光蛋白的文章。 同样看起来是绿色的荧光蛋白,它们的激发和发射光谱、激发效率、斯托克斯位移等等可能完全不一样。 但是有趣的是,光学特性上大相径庭的这些蛋白,不管是蓝的、绿的、黄的、红的,甚至包括那些五颜六色的色蛋白,它们在化学结构、组成上却极为类似。[1]。
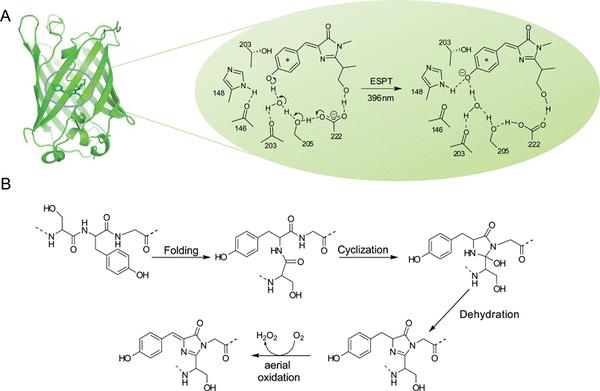
它们基本上都由约230个氨基酸组成,可以说是很小的蛋白质分子了。 结构上我们可以认为荧光蛋白的结构主要由两部分组成,蛋白质骨架和其发色基团,蛋白质骨架基本上都是11链β桶结构,这十一条链交织在一起形成了一个圆桶状的结构(下图)。
通过将发色团与周围的水分子隔离,蛋白质骨架有助于防止在激发态下可能发生的碰撞猝灭。 同时,一些发色基团在蛋白质疏水核心内部的深处也使它免受环境危害,例如卤化物和水合氢离子,这些离子往往会通过改变其离子化状态而降低总荧光。 还有我们的老朋友活性氧(ROS),也有破坏发色团(即被光漂白)的能力,蛋白质骨架也能够保护发色团不受ROS损害。[2]
A 著名的维多利亚水母的GFP蛋白的β桶状结构示意图,右侧放大图的是发色团在受到370-400 nm光线照射时的激发态异构体发出绿色荧光(右)。 B 是发色基团的形成机理
除了蛋白质骨架以外,就是发色团了,发色团的分子量也很小,结构也不是非常复杂。 而且发色团结构的细微改变就能够使得荧光蛋白的荧光特性完全不同,可以看下图,发色基团结构上的非常细微的变化就可以使得荧光蛋白的颜色从蓝色到青色到绿色到黄色到橙色到红色。
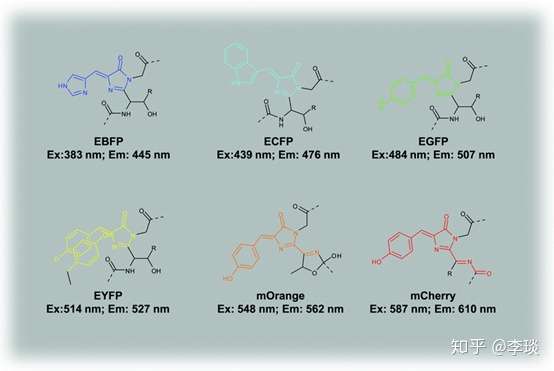
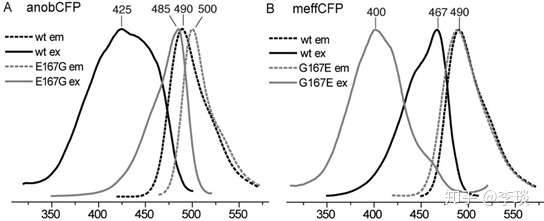
再看这这个图(下图),仅仅一个氨基酸的差别,把两种荧光蛋白的第167个氨基酸替代成谷氨酸以后,A图的青色荧光蛋白吸收峰和发射峰都发生了很大的偏移,吸收峰从425偏到了485nm,发射峰从490变成了500nm,变得更绿了。 而B图的青色荧光蛋白发射峰倒是基本没变,吸收峰从吸收蓝光变成了吸收紫外线。
除了荧光蛋白分子本身的组成结构会影响以外,荧光蛋白分子所处的微环境中的一些物质比如一些小分子的多肽还有一些氨基酸分子等等(别想了,这里面不包括鱼友觉得有用的那些金属离子)对于成熟发色团的形成以及适当的功能也是必不可少的。发色团周围的氨基酸在成熟过程中通过稳定催化中涉及的几个官能团发挥关键作用(别瞎想了,人工加进去的氨基酸也不太可能会对荧光蛋白产生什么影响)。
除此之外,荧光蛋白还有一些非常有趣的光学现象 比如一些荧光蛋白是光诱导的,比如下图A中的PA-GFP,在初始状态下是没有荧光的,但是在紫外线照射后,它的发色团就会发生氧化脱羧,不可逆的转化成绿色荧光蛋白。 下图B中的发色团各位鱼友一定见过,这个Kaede的红色发色基团就是从八字脑(巴厘脑)中分离出来的,你平常看到的八字脑的红色就是这种基团带来的。 这个基团在黑暗中生成的时候是绿色荧光的,但是在400nm波长的紫外照过以后π共轭体系会发生一些变化使得其产生红色荧光,同样的这种变化是不可逆的。 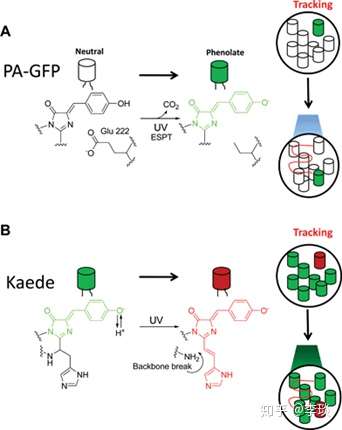
还有一些荧光蛋白的变化是可逆的,可以通过某些手段像开关一样打开关闭。
比如像这个Dronpa发色团,这个发色团可以形成顺反异构两种同分异构体(还记得中学化学学的同分异构体,顺反异构这些东西吧)。 当分子处于顺式构型的时候可以发出绿色的荧光,当强烈的蓝光(488 nm)照射后该基团会变成反式构型的异构体变成为非荧光的“关闭状态”(灰色)。 那么要如何打开这个开关呢,只要将这个发色基团暴露在低水平的紫外线(405Nm)下,基团又可以从非荧光的反式构型重新便会有绿色荧光的顺式构型。 你看,是不是很神奇。 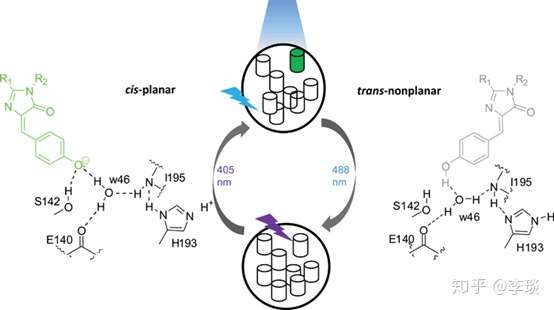
所以你们看,看起来颜色大相径庭的荧光蛋白,甚至是色蛋白,其实他们的结构组成都非常类似,都是一个非常类似的蛋白质骨骼加上看起来大同小异的发色基团组成的。 那么他们为什么那么类似呢?这是因为这些荧光蛋白、色蛋白具有高度的同源性。 这是什么意思呢,可以看看下面这张用各个荧光蛋白克隆出来的序列做的系统发育树 [3] 。 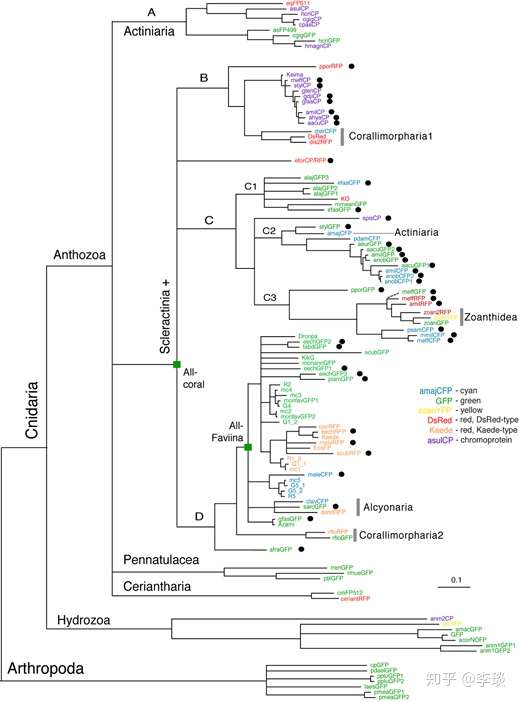 这个系统发育树上包含的这些荧光蛋白大致上可以说明,各种荧光蛋白,包括色蛋白(就是紫色的那几个),其遗传上的差距都不是很远,各个荧光蛋白在系统发育树上所处的位置也与他们所属的shanhu在shanhu的系统进化树上高度类似(除了乱入的那个Zoanthidea),说明了荧光蛋白也是随着shanhu一起进化的。 而且这些荧光蛋白、色蛋白具有共同的祖先,这个最早的绿色荧光蛋白目前我们已经知道最晚从侏罗纪开始就已经出现了。从已知序列反推回去的数据显示最原始的那种荧光蛋白是一种绿色荧光蛋白,最大发射峰为505-506 nm(右图),激发峰值在493-495 nm处(左图) 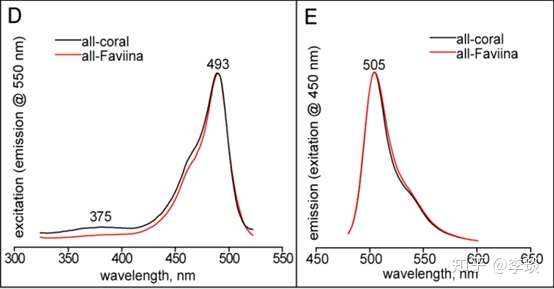
反推预测出来的荧光蛋白祖先序列跟现在的各种荧光蛋白、色蛋白之间序列差别8–12%左右,相似性还是很高的。 也就是说现在的所有荧光蛋白、色蛋白都是从一种绿色荧光蛋白进化来的,那么shanhu为什么要让自己变得五颜六色呢, 目前已经有研究表明shanhu进化出这么多乱七八糟颜色的蛋白是受环境压力选择产生的[4],那么到底是什么样的环境压力导致的呢? 请听下回分解。
还有那个鱼友们最关心的问题,这些结构、组成跟鱼友觉得有用的金属离子可以说是半毛钱关系都没有,所以哪里来的从分子层面影响荧光蛋白呢?这个谣言可以休矣。
参考文献: [1] Lukyanov K A, Chudakov D M, Fradkov A F, et al. Discovery and properties of GFP-like proteins from nonbioluminescent anthozoa.[J]. Methods of biochemical analysis, 2005: 121-138. [2] Sample V, Newman R H, Zhang J, et al. The structure and function of fluorescent proteins[J]. Chemical Society Reviews, 2009, 38(10): 2852-2864. [3] Alieva N O, Konzen K A, Field S F, et al. Diversity and evolution of coral fluorescent proteins.[J]. PLOS ONE, 2008, 3(7): 1-12. [4] Field S F, Bulina M Y, Kelmanson I V, et al. Adaptive Evolution of Multicolored Fluorescent Proteins in Reef-Building Corals[J]. Journal of Molecular Evolution, 2006, 62(3): 332-339.
|  |Archiver|手机版|小黑屋|海友网CMF
( 琼ICP备12003277号 )
|Archiver|手机版|小黑屋|海友网CMF
( 琼ICP备12003277号 )




 发表于 2020-8-9 20:41:25
发表于 2020-8-9 20:41:25
























 发表于 2020-8-9 21:40:23
发表于 2020-8-9 21:40:23

 楼主
楼主