结论珊 瑚礁被描述为海洋沙漠中生物生产力的绿洲(Capone,1992)。周围的贫营养(= 营养枯竭)水域为珊 瑚礁生物群提供的食物很少。因此,复杂的养分循环网已经在珊 瑚礁上形成,以保留和再利用氮和碳等基本元素。珊 瑚礁上的碳循环是多方面的,并且开始被解开。 溶解有机碳(DOC) 水平的可靠测量对于实现这一目标至关重要,岛津 TOC 分析仪的推出代表着研究能力的明确飞跃。在繁荣的珊 瑚礁上,大部分碳输入来自大气中的 CO 2,它首先通过光合作用“固定”成碳水化合物,然后变成许多其他有机分子。珊 瑚中的虫黄藻产生的大部分碳以珊 瑚粘液的形式重新释放。这种珊 瑚的贡献显着丰富了 DOC 库。 该 DOC 是水柱
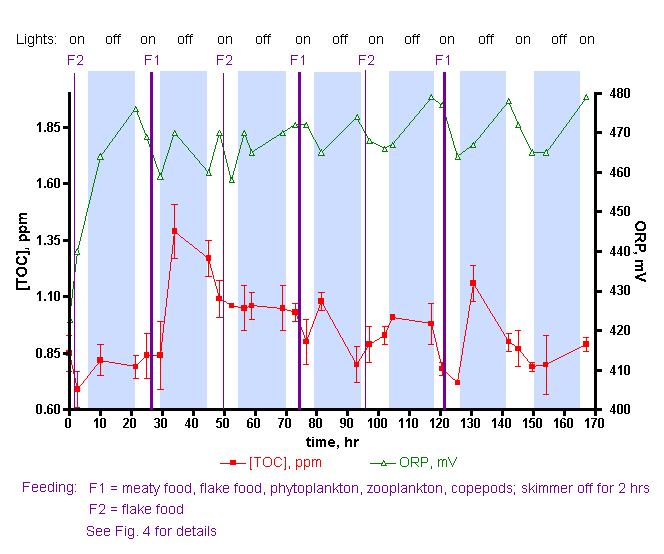
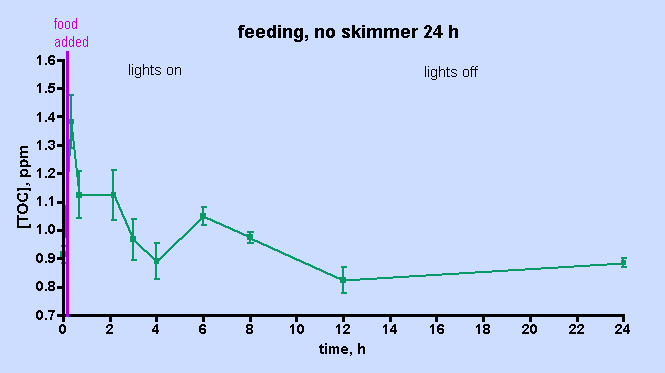
中各种珊 瑚礁微生物群(包括细菌)的主要食物在珊 瑚本身和微型浮游生物等中。最后,这些微生物区系和微型动物作为各种滤食动物的食物,包括珊 瑚。因此,随着碳基营养物质的循环利用,大大小小的珊 瑚礁生物的相互依存关系得以显现。周围的贫营养海洋可以作为缓冲吸收多余的营养物质,否则这些营养物质可能对珊 瑚礁居民有害,并通过进出口调节溶解物种的水平,具体取决于相对(珊 瑚礁与开阔海洋)浓度。我们的圈养珊 瑚礁远远达不到这种养分商业模式;当然,我们没有开阔的海洋缓冲区来稀释废物,因此我们依靠换水来执行这一重要功能,目前尚不清楚我们的水族箱是否包含适当比例的所有成分,促进有效的养分循环所必需的真实珊 瑚礁。尽管如此,我们所做的似乎是有效的,至少在大多数情况下是这样。但是,当事情出错时会发生什么?来自 Rohwer 研究的间接证据表明 DOC 在珊 瑚死亡率中的失衡,失控的细菌生长可能是罪魁祸首。这些观察结果可以为鱼友们提供信息吗?是否有可能改善问题的回应?当然,第一步是在不同的饲养协议下建立健康珊 瑚缸中 TOC (DOC) 水平的基线。在本文中,提供了实现这一目标的数据,并通过与世界各地健康珊 瑚礁的 TOC/DOC 水平进行比较来进一步验证这些 TOC 数字。此外,令人惊讶的最小影响 尽管如此,我们所做的似乎是有效的,至少在大多数情况下是这样。但是,当事情出错时会发生什么?来自 Rohwer 研究的间接证据表明 DOC 在珊 瑚死亡率中的失衡,失控的细菌生长可能是罪魁祸首。这些观察结果可以为鱼友们提供信息吗?是否有可能改善问题的回应?当然,第一步是在不同的饲养协议下建立健康珊 瑚缸中 TOC (DOC) 水平的基线。在本文中,提供了实现这一目标的数据,并通过与世界各地健康珊 瑚礁的 TOC/DOC 水平进行比较来进一步验证这些 TOC 数字。此外,令人惊讶的最小影响 尽管如此,我们所做的似乎是有效的,至少在大多数情况下是这样。但是,当事情出错时会发生什么?来自 Rohwer 研究的间接证据表明 DOC 在珊 瑚死亡率中的失衡,失控的细菌生长可能是罪魁祸首。这些观察结果可以为鱼友们提供信息吗?是否有可能改善问题的回应?当然,第一步是在不同的饲养协议下建立健康珊 瑚缸中 TOC (DOC) 水平的基线。在本文中,提供了实现这一目标的数据,并通过与世界各地健康珊 瑚礁的 TOC/DOC 水平进行比较来进一步验证这些 TOC 数字。此外,令人惊讶的最小影响 当事情出错时会发生什么?来自 Rohwer 研究的间接证据表明 DOC 在珊 瑚死亡率中的失衡,失控的细菌生长可能是罪魁祸首。这些观察结果可以为鱼友们提供信息吗?是否有可能改善问题的回应?当然,第一步是在不同的饲养协议下建立健康珊 瑚缸中 TOC (DOC) 水平的基线。在本文中,提供了实现这一目标的数据,并通过与世界各地健康珊 瑚礁的 TOC/DOC 水平进行比较来进一步验证这些 TOC 数字。此外,令人惊讶的最小影响 当事情出错时会发生什么?来自 Rohwer 研究的间接证据表明 DOC 在珊 瑚死亡率中的失衡,失控的细菌生长可能是罪魁祸首。这些观察结果可以为鱼友们提供信息吗?是否有可能改善问题的回应?当然,第一步是在不同的饲养协议下建立健康珊 瑚缸中 TOC (DOC) 水平的基线。在本文中,提供了实现这一目标的数据,并通过与世界各地健康珊 瑚礁的 TOC/DOC 水平进行比较来进一步验证这些 TOC 数字。此外,令人惊讶的最小影响 是否有可能改善问题的回应?当然,第一步是在不同的饲养协议下建立健康珊 瑚缸中 TOC (DOC) 水平的基线。在本文中,提供了实现这一目标的数据,并通过与世界各地健康珊 瑚礁的 TOC/DOC 水平进行比较来进一步验证这些 TOC 数字。此外,令人惊讶的最小影响 是否有可能改善问题的回应?当然,第一步是在不同的饲养协议下建立健康珊 瑚缸中 TOC (DOC) 水平的基线。在本文中,提供了实现这一目标的数据,并通过与世界各地健康珊 瑚礁的 TOC/DOC 水平进行比较来进一步验证这些 TOC 数字。此外,令人惊讶的最小影响 蛋白质脱脂揭示了 TOC 水平。在这一点上,很明显,如果可以监测 TOC 水平以分析一个撇油器(在本例中为 H&S A200)的影响,那么可以监测它们以测量在实验罐上运行的不同类型的撇油器的影响。此外,这些类型的实验还可用于更直接和定量地探测颗粒活性炭 (GAC) 的 TOC 去除能力,这在 2 号罐与 3 号罐的比较中暗示。这些类型的实验可能第一次允许对撇油器和独立的 GAC 性能进行定量评估,这与似乎围绕这些海洋水族馆维护/设备领域的炒作和错误信息相分离。这些实验正在进行中,结果将在适当的时候报告。 最后,确定阻止珊 瑚缸珊 瑚死亡事件的方法的目标可能受益于从“病态”缸而不是本研究中检查的健康水族箱中收集的数据。如果经历珊 瑚崩溃的坦克在水中的 TOC 含量异常高(或低),但在其他方面具有可接受的水参数,那么一个新的和其他未被重视的恶棍将在手。在这种情况下,鱼友们能做什么呢?Ferrier-Pagès 在Galaxea上描述的一项实验提供了思考的食粮。这种珊 瑚的标本用青霉素、链霉素和两性霉素联合抗生素治疗;随后细菌对 DOC 的吸收停止了,Rohwer 将其确定为过量发生时珊 瑚死亡的机制。
致谢我们感谢宾夕法尼亚州立大学 Eberly 理学院的财政支持,感谢宾夕法尼亚州立大学土木与环境工程系的 Bruce Logan 博士和 David Jones 先生使用岛津 5000 TOC 分析仪,以及 SCARS (州立大学水族馆和珊 瑚礁协会)捐赠水样。
参考
- 安斯沃思,TD;很好,M。罗夫,G。Hoegh-Guldberg, O. 2008。“细菌不是地中海珊 瑚Oculina patagonica漂白的主要原因。” 诠释。社会党。微生物生态学。杂志,2,67-73。
- 阿勒斯,E. 2007a。博士 论文:“海洋浮游细菌对生长条件实验操作的反应”。Fachbereich Biologie/Chemie der Universität Bremen。
- 艾勒斯,E。戈麦斯-康萨尔瑙,L.;平哈西,J。加索尔,JM;西梅克,K。佩恩塔勒,J. 2007b。“ Alteromonadaceae和Rhodobacteriaceae对海洋中宇宙的葡萄糖和磷操纵的反应。” 环境。微生物学,9,2417-2429。
- Baker, AC 2003。“珊 瑚藻共生的灵活性和特异性:共生藻的多样性、生态学和生物地理学。 ” 安努。牧师生态。进化。系统。34, 661-689。
- Benner, R. 2002. 海洋溶解有机物的生物地球化学,学术出版社,圣地亚哥,美国。
- 伯恩,总干事;Munn, CB 2005。“与大堡礁珊 瑚Pocillipora damicornis相关的细菌多样性。” 环境。微生物学,7,1162-1174。
- 卡彭,总干事;东南邓纳姆;霍里根,SG;Duguay, LE 1992。“松散珊 瑚礁沉积物中的微生物氮转化。” 三月生态。编。序列号,80,1992。
- 科斯坦萨,R.;D'Arge, R.; 德格鲁特,R.;法伯,S。格拉索,M。汉农,B。林堡,K。纳伊姆,S。奥尼尔,房车;帕鲁埃洛,J。拉斯金,RG;萨顿,P。van den Belt, M. 1997。“世界生态系统服务和自然资本的价值”。自然,387、253-260。
- 隐蔽,JS;Moran, MA 2001。“使用溶解有机碳的高分子量和低分子量部分的河口细菌群落的分子表征。” 水草。微生物。生态学,25,127-139。
- 克罗斯兰,CJ;巴恩斯,DJ;Borowitzka, MA 1980。“鹿角珊 瑚Acropora acuminata的昼夜脂质和粘液生产。” Mar. Biol., 60, 81-90。
- Crossland, CJ 1987。“从珊 瑚Acropora variabilis和Stylopora pistillata中原位释放粘液和 DOC-脂质。” 珊 瑚礁,6, 35-42。
- 达克洛,硬件;Mitchell, R. 1979。“活珊 瑚粘液层中的细菌种群和适应性”。柠檬醇。Oceanogr., 24, 715-725。
- Ducklow, HW 1983。“海洋中细菌的产生和命运”。生物科学,33,494-501。
- Ducklow, HW 2002。海洋溶解有机物的生物地球化学,学术出版社,圣地亚哥,美国。
- Eppley, RW 1980。“估计中央贫营养海洋中的浮游植物生长率”。布鲁克海文症状。生物学,31,231-242。
- Feldman, KS 2006。“我的 Dialyseas 传奇:优点和缺点”。珊 瑚礁饲养,2006-4 期。
- Ferrier-Pagès,C.;加图索,J.-P.;考韦特,G。乔伯特,J。Allemand, D. 1998。“”虫黄藻珊 瑚束状藻释放溶解的有机碳和氮。” 三月生态。编。第 172、265-274 页。
- Ferrier-Pagès,C.;勒克莱尔,N.;乔伯特,J。Pelegr, SP 2000。“珊 瑚渗出物促进微浮游生物和纳米浮游生物的生长”。水草。微生物。生态学,21,203-209。
- 傅,W。吴,Y。孙,L。Zhang, W. 2007。“海洋海绵Hymeniacidon perleve对综合水产养殖系统中总有机碳 (TOC) 的高效生物修复”。生物技术。生物工程,97,1387-1397。
- 加德纳,TA;科特,IM;吉尔,JA;格兰特,A。Watkinson, AR 2003。“加勒比珊 瑚的长期、区域性衰退”。科学,301, 958-960。
- 冈萨雷斯,JM;西莫,R.;马萨纳,R.;隐蔽,JS;卡萨马约尔,EO;佩德罗斯-阿里奥,C.;Moran, MA 2000。“与产生二甲基磺基丙酸盐的北大西洋藻华相关的细菌群落结构。” 应用程序。环境。微生物学,66, 4237-4246。
- 戈特弗里德,M。Roman, MR 1983。“珊 瑚礁浮游动物对珊 瑚粘液碎屑的摄取和掺入。” 三月生物学,72,211-218。
- 格罗萨特,惠普;列沃尔德,F。阿尔盖尔,M。西蒙,M。Brinkhoff, T. 2005。“海洋硅藻物种拥有独特的细菌群落”。环境。微生物学,7, 860-873。
- 哈塔,H。工藤,S。山野,H。库拉诺,N。Kayanne, H. 2002。“白穗珊 瑚礁(日本石垣岛)的有机碳通量”。三月生态。编。序列号,232、129-140。
- Hedges, JI 2002。海洋溶解有机物的生物地球化学,学术出版社,圣地亚哥,美国。
- 赫森,做;Anderson, TR 2008。“水生生物和生态系统中的过量碳:生理、生态和进化意义。” 柠檬醇。Oceanogr., 53, 1685-1696。
- Holmes-Farley, R. 2004。“珊 瑚礁水族馆中的有机化合物”。珊 瑚礁饲养,http://www.reefkeeping.com/issues/2004-10/rhf/。
- Johannes, RE 1967。“珊 瑚礁附近有机聚集体的生态学”。柠檬醇。海洋学,12, 189-195。
- Kirchman, DL 1990。“亚北极太平洋溶解有机物对细菌生长的限制”。三月生态。编。序列号,62, 47-54。
- 克劳斯,JS;詹斯,我。海库普,JM;桑福德,RA;Fouke, BW 2007。“沿海水深度和沿海污染梯度的珊 瑚微生物群落、虫黄藻和粘液”。环境。微生物学,9,1291-1305。
- 克莱恩,DI;昆茨,新墨西哥州;布赖特巴特,M。诺尔顿,N。Rohwer, F. 2006。“有机碳水平升高和微生物活动在珊 瑚死亡率中的作用。三月生态。编。序列号,314、119-125。
- 库珀曼,N。本-多夫,E.;克拉马尔斯基-温特,E.;巴拉克,Z。Kushmaro, A. 2007。“自然和水族馆环境中与珊 瑚粘液相关的细菌群落”。美联储。欧元。微生物。社会,276,106-113。
- 昆茨,新墨西哥州;克莱恩,DI;桑丁,SA;Rohwer, F. 2005。“”三种加勒比珊 瑚物种的有机碳和营养压力源引起的病理和死亡率。”三月生态。编。序列号,294、173-180。
- 拉婕斯,TC 2005a。“中新世-上新世过渡以来大西洋和印度太平洋共生甲藻的“物种”辐射。” 摩尔。生物。进化,22,570-581。
- 拉杰内斯,TC;桑希尔,DJ;考克斯,英孚;斯坦顿,FG;菲特,WK;施密特,GW 2005b。“在夏威夷珊 瑚礁群落的共生甲藻中观察到高度多样性和宿主特异性。” 珊 瑚礁, 23, 596-603。
- 兰伯特,Y。凯尔曼,D。杜宾斯基,Z。尼赞,Y。Hill, RT 2006。“”红海珊 瑚 真菌scutaria 粘液中可培养细菌的多样性。” FEMS 微生物。生态学,58,99-108。
- Liess, A. 2006. 博士 论文:“底栖食物网中的营养化学计量——藻类、食草动物和鱼类之间的相互作用。” 乌普萨拉大学科学技术学院。
- 手段,JC;Sigleo, AC 1986。“珊 瑚礁粘液对牙买加愉景湾附近胶体池的贡献。” 威牛。三月科学,39,110-118。
- 米克尔,P。理查兹,GN;Yellowlees, D. 1988。“六种珊 瑚粘液的结构研究”。三月生物学,99,187-193。
- 莫里亚蒂,DJW;波拉德,PC;Hunt, WG 1985。“珊 瑚礁水柱中细菌生产的时空变化”。三月生物学,85,285-292。
- 小川,H。Ogura, N. 1992。“测量海水中溶解有机碳的两种方法的比较”。自然,356、696-698。
- 佩吉斯,J。托雷顿,J.-P.;Sempéré, R. 1997。“珊 瑚礁泻湖中的溶解有机碳,采用高温催化氧化和紫外光谱法。” CR学院。科学。巴黎, série IIa, 324, 915-922。
- 平哈西,J。萨拉,MM;哈夫斯库姆,H。彼得斯,F。瓜达约尔,O。马利茨,A。Marrase, C. 2004。“不同浮游植物方案下浮游细菌组成的变化”。应用程序。环境。微生物学,70, 6753-6766。
- 丰富,JH;达克洛,硬件;Kirchman, DL 1996。“赤道太平洋沿线 140 o W 中性单糖的浓度和摄取:葡萄糖对异养细菌活动和 DOM 通量的贡献”。柠檬醇。Oceanogr., 41, 595-604。
- 里奇,A。Lewis, TD 2008。“在三种夏威夷珊 瑚物种中发现的表面相关微生物的细菌组成:Porities compressa、Porities lobata和Montipora capitata。” J. Young Invest.,18,第 6 期。
- 罗威尔,F。布赖特巴特,M。贾拉,J。阿扎姆,F。Knowlton, N. 2001。“与加勒比珊 瑚Montastraea franksi相关的细菌多样性。” 珊 瑚礁, 20, 85-95。
- 罗威尔,F。塞古里坦,V。阿扎姆,F。Knowlton, N. 2002。“珊 瑚相关细菌的多样性和分布”。三月生态。编。序列号,243,1-10。
- 罗森伯格,E。;科伦,O。; 雷舍夫,L。埃弗罗尼,R。Zilber-Rosenberg, I. 2007a。“”微生物在珊 瑚健康、疾病和进化中的作用。” 自然评论:微生物。5,355-362。
- 罗森伯格,E。加利福尼亚州凯洛格;Rohwer, F. 2007b。“珊 瑚微生物学。” 海洋学, 20, 146-154。
- 沙龙,G。Rosenberg, E. 2008。“珊 瑚粘液上的细菌生长”。当前。微生物学,56, 481-488。
- 夏普,JH;本纳,R。贝内特,L。加利福尼亚州卡尔森;道,R。Fitzwater, SE 1993。“海水中溶解有机碳的高温燃烧和化学氧化测量的重新评估”。柠檬醇。海洋学家。38, 1774-1782。
- 夏普,JH;本纳,R。贝内特,L。加利福尼亚州卡尔森;菲茨沃特,东南;佩尔策,ET;Tupas, LM 1995。“海水中溶解有机碳的分析:JGOFS EqPac 方法比较”。三月化学。48、91-108。
- Sharp, JH 2002。海洋溶解有机物的生物地球化学,学术出版社,圣地亚哥,美国。
- 索罗金,易(主编)。1993. 珊 瑚礁生态学。施普林格出版社,柏林。
- 斯特纳,RW;Elser, JJ 2002。生态化学计量:从分子到生物圈的元素生物学。新泽西州普林斯顿出版社。
- 托雷顿,J.-P。1999.“大星盘礁泻湖(斐济)中浮游细菌的生物量、生产和异养活动。” 珊 瑚礁,18, 43-53。
- 范杜伊尔,FC;加斯特,G.|J。2001 “DOC、无机养分和浮游细菌生长的小尺度空间变化与不同珊 瑚礁水类型的联系。” 水生微生物。生态学,24, 17-26。
- 韦斯,M。Simon, M. 1999。“”湖沼浮游细菌消耗不稳定的溶解有机物:氨基酸和碳水化合物的相对意义。” 水草。微生物。生态学,17,1-12。
- 野生的,C。休特尔,M。克鲁特,A。克雷姆,SG;拉希德,MYM;Jergensen, BB 2004。“珊 瑚粘液在珊 瑚礁生态系统中充当能量载体和粒子捕集器。” 自然,428,66-70。
- 耶赫尔,G。夏普,JH;多米尼克,M。海斯,C.;Genin, A. 2003。“共生轴承海绵Theonella swinhoei的原位进料和元素去除:散装 DOC 是碳的主要来源。” 柠檬醇。海洋学家。48、141-149。
- 吉永,我。深见,K。Ishida, Y. 1991。“热带海洋珊 瑚礁水域和远洋水域之间细菌组合 DNA 和蛋白质合成率的比较”。三月生态。编。序列号,76,167-174
|
 [复制链接]
[复制链接]
 |手机版|小黑屋|Archiver|CMF海水观赏鱼网
( 琼ICP备12003277号 )
|手机版|小黑屋|Archiver|CMF海水观赏鱼网
( 琼ICP备12003277号 )


 IP卡
IP卡 狗仔卡
狗仔卡 发表于 2022-9-20 15:19:53
发表于 2022-9-20 15:19:53



 提升卡
提升卡 置顶卡
置顶卡 沉默卡
沉默卡 喧嚣卡
喧嚣卡 变色卡
变色卡 千斤顶
千斤顶 显身卡
显身卡 楼主
楼主



 [/url]
[/url]
 [/url][url=]
[/url][url=] [/url][url=]
[/url][url=] [/url][url=]
[/url][url=] [/url]
[/url] [/url][url=]
[/url][url=] [/url][url=]
[/url][url=] [/url][url=]
[/url][url=] [/url][url=]
[/url][url=] [/url]
[/url]
 [/url][url=]
[/url][url=] [/url][url=]
[/url][url=] [/url][url=]
[/url][url=] [/url]
[/url]





 发表于 2022-9-20 21:27:34
发表于 2022-9-20 21:27:34
